---------------------
Qual o papel da fosfoetanolamina nos seres vivos?
A fosfoetanolamina (PEA) está presente em células de bactérias, protozoários, plantas, fungos e animais. Seu papel ainda não é completamente conhecido. A PEA serve de substrato para a produção de diversos fosfolipídios de membrana como a fosfatidiletanolamina e a fosfatidilcolina (Figs. 1, 2). Em bactérias parasitas, a molécula é adicionada a outras presentes na membrana ajudando os micro-organismos a escaparem do sistema imunológico do hospedeiro (Packlam et al 2014). Diversos efeitos contraditórios têm sido atribuídos à PEA em células animais: como estimulação (Karagezyan & Ovsepyan 1975) e inibição (Modica-Napolitano & Renshaw 2004) da atividade oxidativa das mitocôndrias; a promoção da divisão celular e/ou sobrevivência e crescimento (Kano-Sueoka et al. 1979; Kano-Sueoka & Errick 1981; Kiss 1999) e indução da apoptose celular, como defendido pelo grupo de Chierice (Ferreira et al. 2011, 2013) - porém, há indicativos de que a amostra utilizada apresentava um grau muito baixo de pureza, menos de um terço em massa seria de fato constituído de PEA (Dias et al. 2016); podendo ocorrer aumento (Vermeersch et al.2014) e redução (Cady et al. 2010) de concentração celular/tecidual durante hipóxia (baixo teor de oxigênio)/isquemia (baixa irrigação sanguínea).


A fosfoetanolamina apresenta-se em concentração elevada em vários tecidos durante o desenvolvimento embrionário e fetal: células musculares de frango (Granata et al. 2000); músculo esquelético e cardíaco de ratos (Turner et al. 1994); cérebro de coelho (Cohen & Lin 1962);cérebro, coração, fígado, rim e baço de camundongos (Kataoka et al. 1991); cérebro de cães (Gyulai et al. 1984); pool de corpo inteiro de embriões da lagosta europeia (Homarus gammarus) (Rosa et al.2005). Também apresenta-se aumentado no útero pós-parto de coelhas (Cawkwell 1958), de ratas e mulheres (Phoenix & Wray 1994). A presença da PEA no leite materno também aumenta com o tempo de lactação (Harzer et al. 1984) e a concentração no colostro de mulher após parto precoce é mais alta do que no colostro de mulher após parto a termo (Pamblanco et al. 1989). Por outro lado, sua concentração em tecido cerebrais de indivíduos com doenças neurodegenerativas (Alzheimer e Huntington) é reduzida (Ellison et al. 1987). (Tabela 1.) Essas observações não parecem ser facilmente explicadas pelo efeito apoptótico defendido pelo grupo de Chierice; mas são compatíveis com o efeito mitogênico proposto por Kano Sueoka e colaboradores.
Tabela 1. Concentrações de fosfoetanolamina em diferentes tecidos e órgãos. Em itálico: tecidos tumorais.
pu - peso úmido de tecido. Fontes: Ackerstaff et al. 2003; Bell & Bhakoo 1998, Ellison et al. 1987, Granata et al. 2000, Gribbestad et al. 1999, Podo 1999.
Tecido/Órgão
|
Concentração (μmol/g pu)
|
humano
lobo frontal (feto 5 meses)
lobo frontal (feto 8 meses)
lobo frontal (adulto 19 anos)
córtex frontal (adulto 49,8±4,6 anos)
córtex frontal (adulto 70,6±5,0 anos)
córtex frontal (adulto Alzheimer)
córtex frontal (adulto Huntington)
lobo occipital (feto 5 meses)
córtex occipital (adulto 49,8±4,6 anos)
córtex occipital (adulto 70,6±5,0 anos)
córtex occipital (adulto Alzheimer)
córtex occipital (adulto Huntington)
lobo temporal (feto 8 meses)
córtex temporal (adulto 70,6±5,0 anos)
córtex temporal (adulto Alzheimer)
núcleo accumbens (adulto 49,8±4,6 anos)
núcleo accumbens (adulto Huntington)
núcleo caudado (feto 5 meses)
núcleo caudado (feto 8 meses)
núcleo caudado (adulto 19 anos)
caudado (adulto 49,8±4,6 anos)
caudado (adulto Huntington)
hipotálamo (feto 5 meses)
hipotálamo (feto 8 meses)
hipotálamo (adulto 19 anos)
hipocampo (adulto 70,6±5,0 anos)
hipocampo (adulto Alzheimer)
putâmen (adulto 49,8±4,6 anos)
putâmen (adulto Huntington)
córtex cerebral
astrocitoma
astrocitoma anaplástico
glioblastoma
ependimoma
meduloblastoma
meningioma
neurinoma
craniofaringioma
cordoma
adenoma da pituitária
linfoma maligno
adenocarcinoma pulmonar
mama
tumor mamário
fígado
carcinoma hepatocelular
|
7,43
6,63
0,71
0,81±0,11
0,97±0,12
0,50±0,12
0,72±0,06
5,24
0,75±0,17
0,90±0,18
0,80±0,19
0,66±0,10
6,90
1,21±0,14
0,43±0,10
1,25±0,19
0,65±0,10
9,29
8,45
1,04
1,02±0,19
0,25±0,03
6,68
3,82
0,38
0,95±0,13
0,57±0,09
1,18±0,15
0,56±0,06
1,06±0,10
1,27±0,11
2,05±0,41
2,38±0,38
1,06
4,62
1,61±0,24
1,87±0,31
0,68±0,12
2,05±0,69
4,41±1,06
4,29±0,17
2,12±0,20
0,16±0,01
2,1±1,1
0,16±0,10
2,47±0,84
|
rato
útero (feto 20-21 dias)
útero (neonato 1-3 dias)
útero (fêmea adulta grávida)
útero (fêmea adulta pós-parto)
intestino delgado do músculo liso (adulto)
intestino delgado do músculo liso (adulta grávida)
músculo esquelético (feto 20-21 dias)
músculo esquelético (neonato 1-3 dias)
músculo esquelético (adulto)
coração (feto 20-21 dias)
coração (neonato 1-3 dias)
coração (adulto)
cérebro (feto 20-21dias)
cérebro (neonato 1-3 dias)
cérebro (adulto)
|
2,45±0,30
1,65±0,10
1,68±0,14
2,42±0,12
1,63±0,08
2,01±0,16
~1
~1
0,02±0,1
1,2±0,1
1,4±0,1
~0,3
4,13±0,28
3,87±0,55
~1
|
galinha
músculo peitoral (embrião 9 dias)
músculo peitoral (embrião 14 dias)
músculo peitoral (embrião 17 dias)
músculo peitoral (recém eclodido)
|
1,85±0,90
1,33±0,66
0,57±0,33
0,21±0,08
|
A PEA é produzida também pela degradação da esfingosina-1-fosfato (S1P) (Fig. 3). A S1P é um lipídio com vários efeitos no organismo: inflamação, tumorigênese e neovascularização, entre outros (Pyne & Pyne 2010).
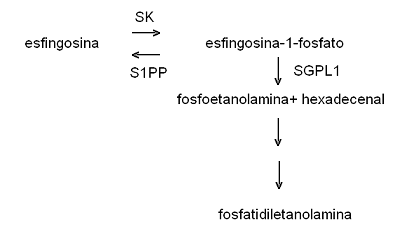
No entanto, células geneticamente modificadas com superexpressão da enzima esfingosina quinase (SK), que converte a esfingosina em S1P não apresentam uma taxa aumentada de divisões celulares. Somente quando a modificação também leva à superexpressão da enzima esfingosina-1-fosfato liase (SGPL1), que quebra a S1P nas moléculas PEA e hexadecenal, o efeito mitogênico é observado, sugerindo, assim, que o produto da ação conjunta das enzimas SK e SGPL1 e não a S1P em si que apresenta o efeito de estimulação da divisão celular. (Kariya et al. 2005.) Sendo a fosfoetanolamina um dos produtos da degradação da S1P, há mais acordo com a proposta da ação mitogênica da PEA, do que de um efeito inibidor ou apoptótico. Por outro lado, a superexpressão S1P fosfatase (S1PP), leva à redução da concentração celular de S1P e aumento da esfingosina e ceramida e induz ao surgimento de características apoptóticas (Mandala et al. 2000).
Em Smith et al. (1984), PEA a 1 μg/ml, na presença de outros fatores como soro fetal e insulina, apresentou efeito mitogênico para células mioepiteliais derivadas de um tumor mamário de rato; porém outras linhagens derivadas de epitélios de tumor mamário e de células mamárias não-tumorais de ratos não foram estimuladas pela PEA. Em Dhakshinamoorthy et al. (2015), PEA a 8 e 12 mM (1.692,76 1 μg/ml) estimulou a apoptose em células Jurkat (linhagem imortalizada derivada de linfócitos T humanos), mas não em linfócitos normais - apresentando até um possível efeito protetivo sobre estas últimas. Kano-Sueoka & Errick (1981), usando linhagens hormônio-dependentes de carcinoma mamário de rato expostas a uma concentração de 5 nmol/ml (0,71 μg/ml) de PEA, obtiveram um aumento de 3,4 vezes na contagem de células em relação ao controle. A mesma linhagem havia sido testada em Kano-Sueoka et al. (1979): o efeito apresentou uma relação dose dependente, aumentando até a concentração de 5 nmol/ml, na concentração de cerca de 50 nmol/ml (7,05 μg/ml), o efeito foi similar à concentração de 5 nmol/ml; em Kano-Sueoka & Errick. (1981), o efeito da PEA foi sempre crescente até a uma concentração de 10 nmol/ml (1,41 μg/ml). A linhagem hormônio-independente de carcinoma mamário de rato não apresentou a mesma resposta clara; células de hepatoma de ratos, células 3T3 de camundongos e células de fibroblasto de ratos não tiveram sua taxa de divisão aumentada pela PEA; células epiteliais normais de ratos, em testes preliminares dos autores, respondem à molécula (Kano-Sueoka et al. 1979). A etanolamina, molécula precursora da fosfoetanolamina, em Kano-Sueoka & Errick (1981), também apresentou efeito mitogênico sobre as células de carcinoma mamário de rato; mas em concentrações maiores (50 nmol/ml), apresentou efeito inibidor do crescimento. Os efeitos mitogênico e apoptótico da PEA parecem ser variáveis de acordo com a linhagem celular e a concentração aplicada.
Em células de câncer, a PEA também apresenta-se em concentrações elevadas, de duas a vinte vezes, bem como a fosfocolina (PCh) (Podo 1999) (Tabela 1). Células de câncer mamário humano triplo negativas (com três mutações que interrompem a atividade de três genes) MDA-MB-231 tratadas com ARNs interferentes pequenos (siRNA, small interfering RNA) que inibem a transcrição de quinase de etanolamina-1 (EtnK-1) e da quinase de colina-α (ChK-α) levam à redução das concentrações intracelulares de PEA e PCh. A aplicação de siRNA ChK-α também provocou a redução da viabilidade das células em 60%; o siRNA EtnK-1 reduziu a viabilidade celular em 50%. (Shah et al. 2015.) A PEA, assim como a PCh, parecem ter um papel importante na viabilidade de células tumorais, mas em desacordo com a proposta do grupo de Chierice.
A concentração intracelular de PEA aumenta ligeira, mas consistentemente, com perturbações no metabolismo celular de naturezas diversas: privação de glicose, condição de hipóxia e de isquemia (Vermeersch et al. 2014; cf. Cady et al. 2010). Isso indica que essa elevação faça parte de um mecanismo genérico de resposta ao estresse. Ela pode resultar de um aumento na taxa de substituição dos fosfolipídios de membrana ou, se o grupo de Chierice estiver certo, de resposta apoptótica ao estresse. (Vermeersch et al. 2014.)
No cérebro do peixe euritérmico (capaz de se adaptar a uma gama ampliada de temperaturas) Perccottus glenni (frequentemente grafado erroneamente como P. glehni), a concentração de PEA aumenta quando o animal é exposto a baixas temperaturas: tanto sazonalmente, quanto em condição de choque térmico. Possivelmente devido à alteração da composição de lipídios da membrana. (Karanova 2013.)
No processo de aclimatação a baixas temperaturas do caruncho do trigo, Sitophilus granarius, e do besourinho dos grãos, Cryptolestes ferrugineus, ocorre também variação nos níveis de fosfoetanolamina no corpo. Em S. granarius, a concentração de PEA diminui com a aclimatação; já em C. ferrugineus, a concentração de PEA aumenta. (Fields et al. 1998.)
Outros efeitos estudados da fosfoetanolamina são: coagulação sanguínea - tem efeito anticoagulante (Koppel et al. 1959); ação em células do núcleo septal em cérebros de ratos - aumenta a captação de colina e síntese de acetilcolina (Bostwick et al. 1992); em neurônios do hipocampo de ratos - aumenta temporariamente a excitabilidade (Zeise & Lehmann 1987).
Conclusão Até o presente, basicamente o único papel bem estabelecido da fosfoetanolamina nos organismos é como precursor de importantes fosfolipídios de membrana celular. Vários outros interessantes efeitos têm sido atribuído à molécula: regulação do ciclo celular, ação sobre o metabolismo oxidativo, resposta ao estresse... Mas, como boa parte desses efeitos parecem variar e até apresentar resultados contrários entre diferentes estudos, seria necessário um esforço de replicação independente dos testes realizados até o momento; além de ampliação das pesquisas para determinar os fatores responsáveis por tal variação. Certamente outras funções biológicas da PEA deverão ainda ser descobertas. A maioria dos indícios, no entanto, parecem indicar que a PEA não tem efeito inibidor sobre células tumorais em concentrações fisiológicas normais. Apenas em concentrações de 10 a 100 vezes acima da normal parecem produzir efeitos de inibição ou de apoptose em certas células cancerosas e mesmo tal efeito é incerto pela possível contaminação da PEA utilizada nesses estudos do grupo de Chierice.
Referências
Ackerstaff, E. et al. 2003. A target in cancer cells? Journal of Cellular Biochemistry 90: 525-33.
Bell, J.D. & Bhakoo, K.K. 1998. Metabolic changes underlying 31P MR spectral alterations in human hepatic tumours. NMR in Biomedicine 11(7): 354-9.
Bostwick, J.R. et al. 1992. Phosphoethanolamine enhances high-affinity choline uptake and acetylcholine synthesis in dissociated cell cultures of the rat septal nucleus. Journal of Neurochemistry 59(1): 236–44.
Cady, E.B. et al. 2010. Coupling of cerebral phosphoethanolamine and nucleotide triphosphate levels and mitochondrial-respiration modulation during perinatal 'secondary energy failure'. Proc. Intl. Soc. Mag. Reson. Med. 18: 2395.
Cawkwell, J.M. 1958. Phosphate esters in the uterus. Biochem. Journal 70(2): 248-52.
Cohen, M.D. & Lin, S. 1962. Acid soluble phosphates in the developing rabbit brain. Journal of Neurochemistry 9: 345-52.
Dhakshinamoorthy, S. et al. 2015. Metabolomics identifies the intersection of phosphoethanolamine with menaquinonetriggered apoptosis in an in vitro model of leukemia. Mol. BioSyst. 11: 2406-16.
Dias, L,C. et al. 2016. Caracterização do conteúdo das cápsulas de fosfoetanolamna (FOS) para o MCTI: análise de ressonância magnética nuclear de 1H, 13C e 31P. Retório técnico-científico. LQOS/Unicamp. Campinas, SP. Disponível em: http://www.mctic.gov.br/mctic/export/sites/institucional/ciencia/SEPED/Saude/fosfoetanolamina/arquivos/Relatorio-Completo-de-Sintese-e-Caracterizacao.pdf. Acessado em: 26.jan.2018.
Ellison, D.W. et al. 1987. Phosphoethanolamine and ethanolamine are decreased in Alzheimer's disease and Huntington's disease. Brain Research 417: 389-92.
Ferreira, A.K. et al. 2011. Synthetic phosphoethanolamine induces apoptosis through caspase-3 pathways by decreasing expression of Bax/Bad protein and chanves cell cycle in melanoma. J. Cancer Sci. Ther. 3: 53-9.
Ferreira, A.K. et al. 2013. Synthetic phosphoethanolamine induces cell cycle arrest and apoptosis in human breast cancer MCF-7 cells through the mitochondrial pathway. Biomed. Phamarcother. 67: 481-7.
Fields, P.G. et al. 1998. The effect of cold acclimation and deacclimation on cold tolerance, trehalose and free amino acid levels in Sitophilus granarius and Cryptolestes ferrugineus (Coleoptera). Journal of Insect Physiology 44: 955-65.
Granata, F. et al. 2000. Phosphocoline and phosphoethanolamine during chick embryo myogenesis> a 31P-NMR study. Biochimica et Biophysica Acta 1483: 334-42.
Gribbestad I.S. et al.1999. Metabolite composition in breast tumors examined by proton nuclear magnetic resonance spectroscopy. Anticancer Res 19: 1737–46.
Gyulai, L. et al. 1984. Phosphorylethanolamine - the major constituent of the phosphomonoester peak observed by 13P-NMR on developing dog brain. FEBS 178(1): 137-42.
Harzer, G. et al. 1984. Human milk nonprotein nitrogen components: changing patterns of free amino acids and urea in the course of early lactation. The American Journal of Clinical Nutrition 40: 303-9.
Kano-Sueoka, T. et al. 1979. Phosphoethanolamine as a growth factor of mammary carcinoma cell line of rat. Proc. Natl. Acad. Sci. 76: 5741-4.
Kano-Sueoka, T. & Errick, J.E. 1981. Effects of phosphoethanolamine and ethanolamine on growth of mammary carcinoma cells in culture. Experimental Cell Reserach 136: 137-45.
Karanova, M.V. 2013. Phosphoethanolamine in brain of the euritherm lake fish Perccottus glehni (Eleotridae, Perciformes, Dyb. 1877) as a phenomenon depending on temperature factor. Journal of Evolutionary Biochemistry and Physiology 49(3): 291-9.
Karagezyan, K.G. & Ovsepyan, L.M. 1975. [Role of ethanolamine, phosphoethanolamine, and phosphatidylethanolamine in oxidative phosphorylation of albino rat brain mitochondria.] Byulleten' Éksperimental'noi Biologii i Meditsiny 80(10):58-9.
Kariya, Y. et al. 2005. Products by the sphingosine kinase/sphingosine 1-phosphate (S1P) lyase pathway but not S1P stimulate mitogenesis. Genes to Cells 10: 605–15.
Kataoka, H. et al. 1991.O-phosphoethanolamine content in mouse tissues during development. Agric. Biol. Chem. 55(1): 289-90.
Kiss, Z. 1999. Regulation of mitogenesis by water-soluble phospholipid intermediates. Cell. Signal 11(3): 149-57.
Koppel, J.L. et al. 1959. Effects of phosphoethanolamine and phosphoserine on blood coagulation mechanism. American Journal of Physiology 196: 1020-4.
Mandala, S.M. et al. 2000. Molecular cloning and characterization of a lipid phosphohydrolase that degrades sphingosine-1-phosphate and induces cell death. PNAS 97: 7859–64.
Modica-Napolitano, J.S. & Renshaw, P.F. 2004. Ethanolamine and phosphoethanolamine inhibit mitochondrial function in vitro: implications for mitochondrial dysfuncion hypothesis in depression and bipolar disorder. Biol. Psychiatry 55: 273-7.
Mudd, S.H. & Datko, A.H. 1986. Phosphoethanolamine bases as intermediates in phosphatidylcholine synthesis by Lemna. Plant Physiol. 82: 126-35.
Pamblanco, M. et al. 1989. Free amino acids in preterm and term milk from mothers delivering appropriate- or small-for-gestational-age infants. Am. J. Clin. Nutr. 50: 778-81.
Phoenix, J. & Wray, S. 1994. Changes in human and rat uterine phosphoethanolamine and taurine with pregnancy and parturition. Experimental Physiology 79: 601-4.
Plackam, M. et al. 2014. Phosphoethanolamine decoration of Neisseria gonorrhoeae lipid A plays a dual immunostimulatory and protective role during experimental genital tract infection. Infection and Immunity 82(6): 2170-9.
Podo, F. 1999. Tumour phospholipid metabolism. NMR in Biomedicine 12(7): 413-39.
Pyne, N.J. & Pyne, S. 2010. Sphingosine-1-phosphate and cancer. Nature Reviews Cancer 10, 489-503.
Rosa, R. et al. 2005. Changes in amino acids and lipids during embryogenesis of European lobster, Homarus gammarus (Crustacea: Decapoda). Comparative Biochemistry and Physiology, part B, 140: 241-9.
Shah, T. et al. 2015. Ethanolamine kinase-1 and phosphoethanolamine are potential diagnostic markers and therapeutic targets in breast cancer [abstract]. In: Proceedings of the 106th Annual Meeting of the American Association for Cancer Research; 2015 Apr 18-22; Philadelphia, PA. Philadelphia (PA): AACR; Cancer Res 2015;75(15 Suppl):Abstract nr 1195.
Smith, J.A. et al. 1984. Different growth factors stimulate cell division of rat mammary epithelial, myoepithelial, and stromal cell lines in culture. Journal of Cellular Physiology 119: 320-6.
Spiegel, S. & Milstien, S. 2003. Sphingosine-1-phosphate: an enigmatic signaling lipid. Nature Reviews Molecular Cell Biology 4: 397-407.
Turner, O. et al. 1994. Developmental and gestational changes of phosphoethanolamine and taurine in rat brain, striated and smooth muscle. Experimental Physiology 79: 681-9.
Vance, J.E. 2008. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related aminophospholipids. Journal of Lipid Research 49: 1377-87.
Vance, J.E. & Vance, D.E. 2004. Phospholipid biosynthesis in mammalian cells. Biochem. Cell Biol. 82: 113-28.
Vermeersch, K.A. et al.. 2014. Distinct metabolic responses of an ovarian cancer stem cell line. BMC Systems Biology 8: 134.
Zeise, M. & Lehmann, A. 1987. Phosphoethanolamine transiently enhances excitability of rat hippocampal neurons in vitro. J Comp Physiol A 161: 461-7.


{kind=link}
Nenhum comentário:
Postar um comentário